This is part of my “journal club for credit” series. You can see the other computational neuroscience papers in this post.
Unit: Diffusion
Organized by Ben Regner
- Standard Diffusion
- Anomalous Diffusion
- Life at Low Reynold’s Number
Papers
- Random walk models in biology. By Codling, Plank, and Benhamou in 2008.
Other Useful References
What is anomalous diffusion?
If one measures the mean square displacement vs time, it can be parameterized as

where 
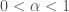

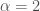
How to describe anomalous diffusion?
Currently, there is no “best” or “simple” description of anomalous diffusion in the general case. However, continuous-time random walks (CTRW) are one paradigm that I find helpful as a conceptual and simulation framework.
In the simplest discrete random walk (DRW), at every time step, a particle makes a jump of fixed size, the only question is the direction. The next generalization has the particle make a jump at every time step, but now it draws the jump size from a distribution.
The idea of a CTRW is that there is now a distribution both of the waiting time between jumps, and the jump size. If the waiting time follows the exponential distribution and the jump size follows the normal distribution, one ends up with the Wiener process aka standard diffusion and Brownian motion.
What causes anomalous diffusion?
1. Increments are independent
2. Increments are wide sense stationary. 1st moment and autocovariance don’t depend on time (this is weaker condition then complete stationarity)
3. Zero mean
The third condition is often ignored by examining the motion relative to the mean displacement (ie the actual displacement is not Brownian, but fluctuations in the displacement could be Brownian). So really, the first two are the more important conditions. Therefore, anomalous diffusion arises due to non-independent increments and/or correlations in time of the mean and/or standard deviation.
The CTRW allows one to think more precisely about different mechanisms that can give rise to anomalous diffusion. There is not one single way to get sub or super-diffusion in CTRW, since there are two, potentially dependent, distributions (waiting time and jump size). However, there are a few common situations that seem to arise often in biology and elsewhere (see Random walk models in biology, Box 2 for original idea). Subdiffusion in biology is often caused by longer waiting time distributions (compared to exponential), or molecular crowding, while superdiffusion in occurs when jump sizes are drawn from a Levy flight or other alpha stable distributions.
Examples
For further exploration of anomalous diffusion in biology, I recommend these papers
Advanced Questions
- This is an interesting paper that introduces a renormalization group approach to classifying diffusion processes

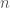 is the number of particles,
is the number of particles,  is the location of the particles,
is the location of the particles,  is the diffusion constant, and
is the diffusion constant, and 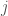 is the flux of particles.
is the flux of particles.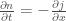

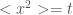
 is superdiffusive, and ballistic is
is superdiffusive, and ballistic is  in certain turbulent regimes.
in certain turbulent regimes.
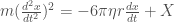
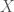 is a stochastic variable. It is assumed to be zero mean, unit variance, and no time correlations, aka
is a stochastic variable. It is assumed to be zero mean, unit variance, and no time correlations, aka 